page26
Note: the contents of this page as well as those which precede and follow, must be read as a continuation and/or overlap in order that the continuity about a relationship to/with the dichotomous arrangement of the idea that one could possibly talk seriously about peace from a different perspective as well as the typical dichotomous assignment of Artificial Intelligence (such as the usage of zeros and ones used in computer programming) ... will not be lost (such as war being frequently used to describe an absence of peace and vice-versa). However, if your mind is prone to being distracted by timed or untimed commercialization (such as that seen in various types of American-based television, radio, news media and magazine publishing... not to mention the average classroom which carries over into the everyday workplace), you may be unable to sustain prolonged exposures to divergent ideas about a singular topic without becoming confused, unless the information is provided in a very simplistic manner.
Let's face it, humanity has a lousy definition, accompanying practice, and analysis of peace.
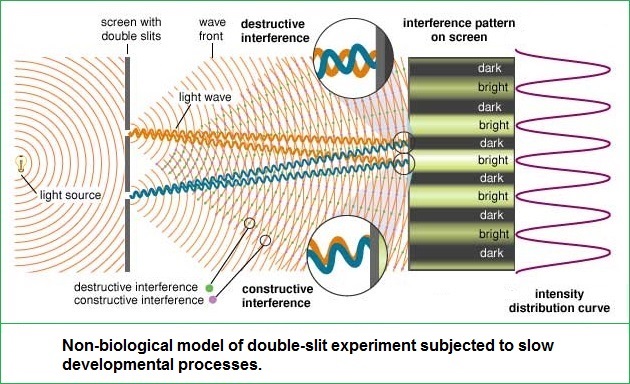
As an extension to the former page, though all pages in this series are linked, let us pursue a more definitive examination of the two-slit experiment with an enlarged philosophy. In this sense, we will recognize that the usage of more-than-two slits does increase the quantity of interference patterns, but that the light/dark bands remains a relevant part of the discussion. Also, it is of some interest to point out that some "three-slit" experiments involved a type of pyramidal configuration in that one slit was placed in front of two slits, as shown in the accompanying image which was adapted from a two-slit model to portray this idea:
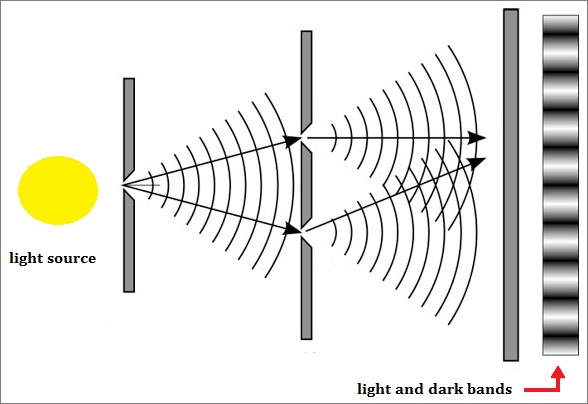
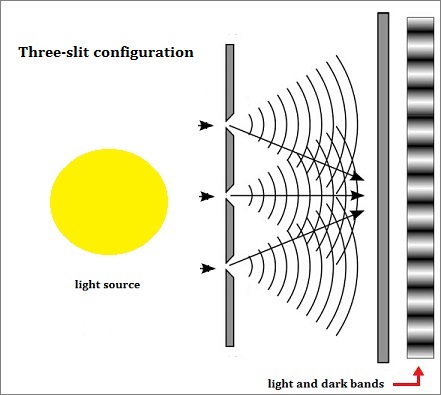
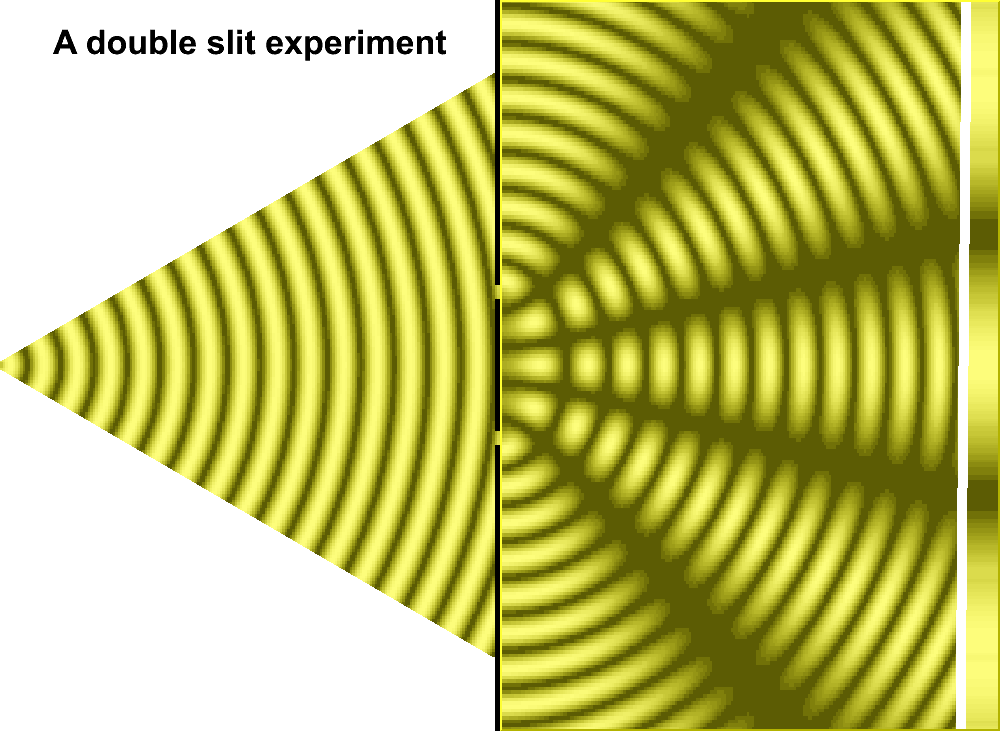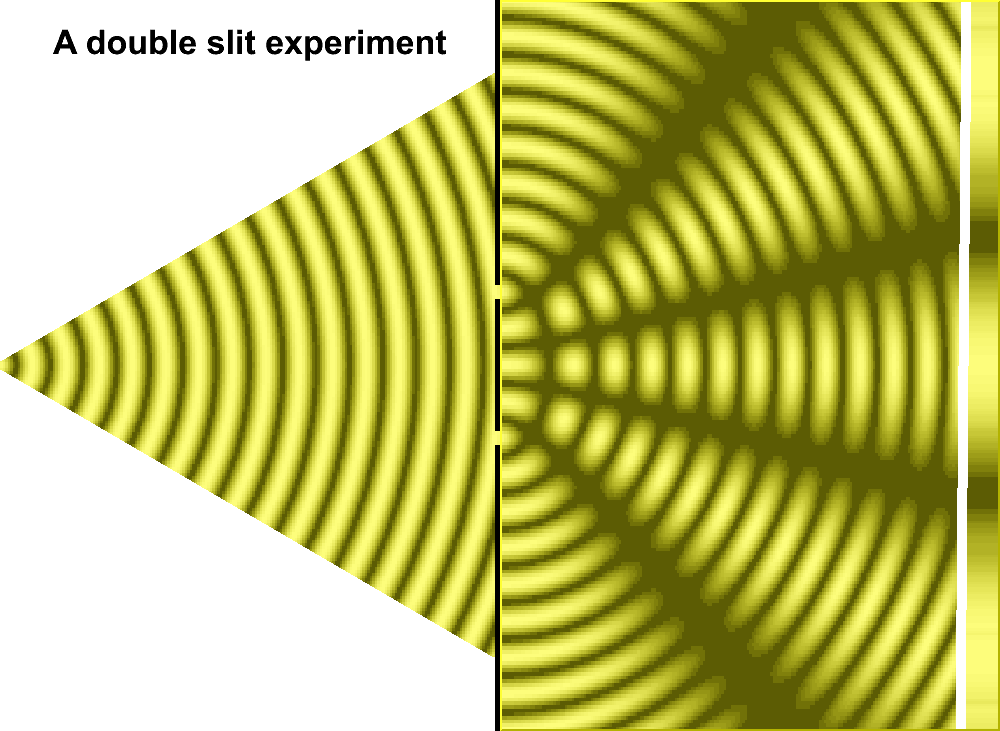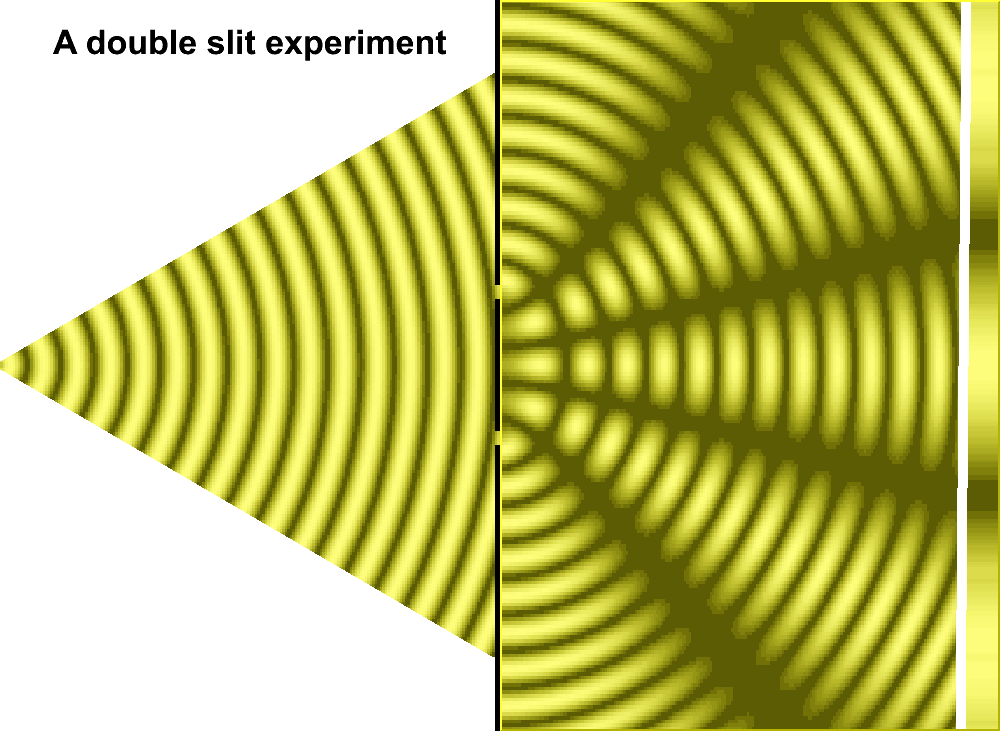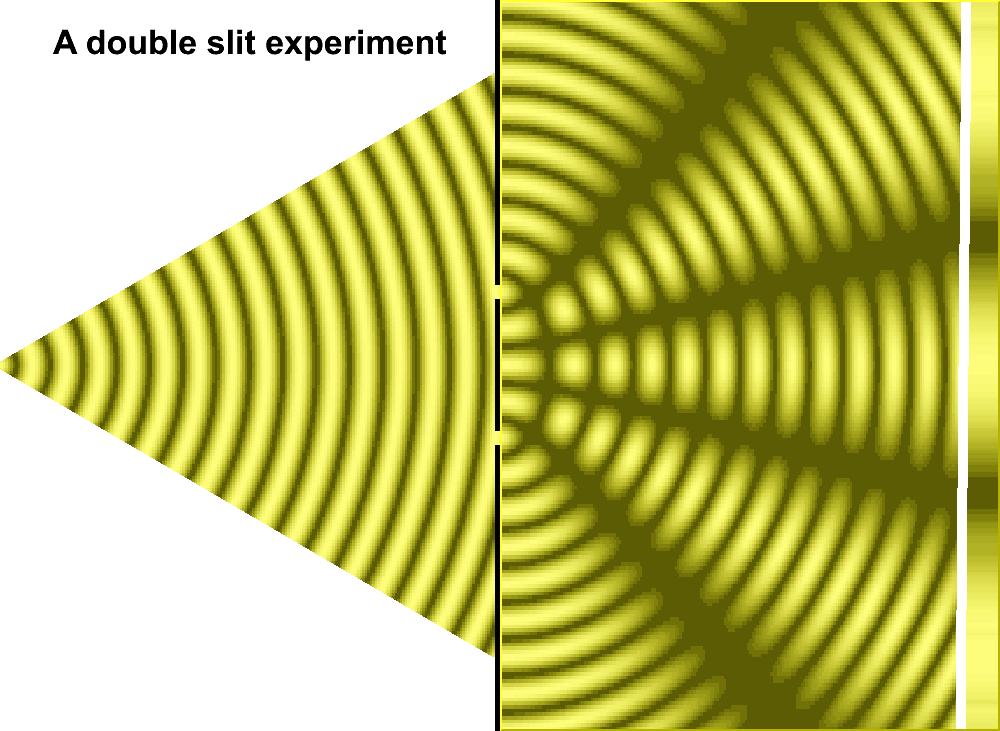
An actual three-slit model might well be illustrated by the image to the right or the following generalized image, so long as one does not try to get too technical with respect to what direction the actual light source may illuminate the slits, how large and how intense the light is, size of the slits, the distance between slits, etc... And as an added feature to assist those whose minds do not typically generate conceptions in an animated form as we may presume Einstein did in his thought experiments, an animated image of a two-slit model is provided:
|
|
What may be overlooked by some readers is that there is a wavering pattern due to light intensity because of the variability of fluctuations inherent in the incoherency of light:
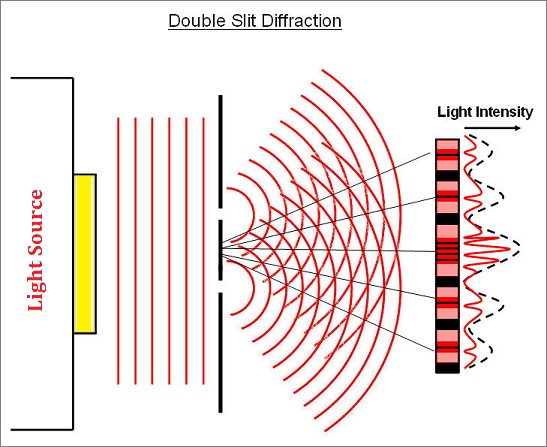 |
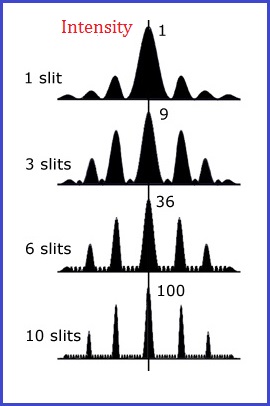 |
The usage of additional quantities of slits would produce different collections of interference patterns, that may be illustrated in a different manner:
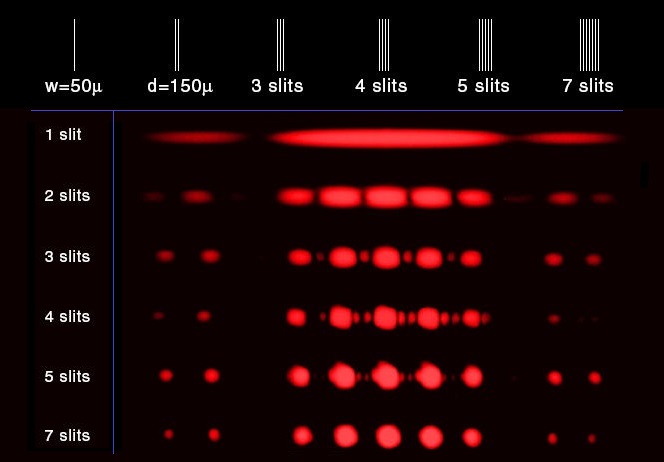
Here's a PBS (Public Broadcasting Station) representation of the double-slit experiment from its Space Time series, which was retrieved from the Popular Mechanics/ Science division page. Because it is a help-us-get-funding effort, it is an embellished entertainment production:
Now let's get a bit more nit picky by questioning the type of hole being used, the distant between the holes, etc.
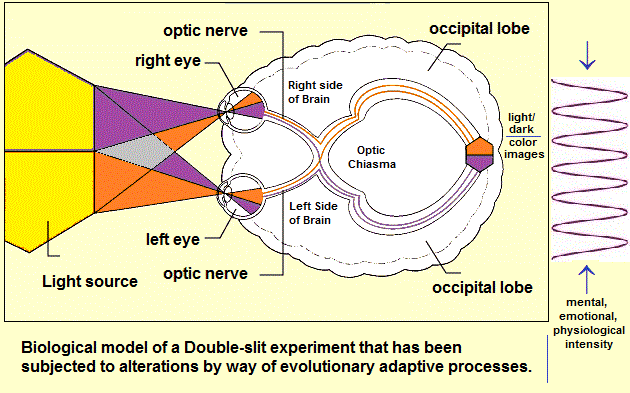
Returning to the previous page's reference that the two-slit model gives the impression of an "anthropomorphism" (has the human attribute of two eyes), let us pursue this idea and ask why a "slit" was used instead of a hole that more clearly represents the human eye called the pupil? And let us add that the spacing between the holes differs in as much as the distant between the eyes of different humans and other animals, including insects, with the "eyes" of plants placed so as to take advantage of (or hide from) the sun, and that many in fact follow the Sun, thus describing that the distant between the "eye(s)" of a plant can be calculated if one cared to do so. In other words, if the "eye(s)" of a plant was wide, some plants wouldn't have to have a sun/moon-orienting system called helio or lunar-tropism... though there are other tropisms as well.
If we say the two slits represent two "slanted" eyes with a particular distance between them and then seek out such a configuration in the animal/insect kingdom, we might not find the exact configuration being used in the two-slit model, thus identifying an artificiality.
 |
 |

While the shape of the human eye is generally considered to be circular, those who have lived for generations in regions with blowing sand and or bright sunlight that did not use some artificially developed means of protecting the eyes, may have developed what is colloquially referred to as the "slanted eye" (almond-shaped) appearance created by the lower and upper eyelids being "stretched" outward; whose inner and outer "points" are called canthi. In other words, the amount of light being permitted to service the whole eye ball is reduced. Another mechanism that is used to reduce or increase the amount of light entering into the eye is the flexibility of the pupil to open or close, as seen in the image to the right that is being illustrated by two different cat eyes. Please note that for the most part, the slit is vertical as is described by the usage of slits in the double-slit experiment. |
|
|
However, if there are no eyelids as in humans, nor a third means of filtering called the nictating membrane in the Tuatara (a lizard-like reptile of the coastal islands of New Zealand), Crocodilians and most birds, and there is no flexible pupil; the changes in "aperture" size (related to usable light entering the eye), must thus rely on some other mechanism such as the mechanism of sight and the human brain to alter the shutter speed, lens magnification, and color lens scheme as seen in photography. (Humans have three color filters.) The shape of the pupil, the rate at which eye lids can open and shut, the distance between the light source and openings as well as the openings themselves, the distance from the opening and the "image catching wall" upon which only shadows may fall to give a representation like the idea in the story of Plato's cave; are all variables to be considered in a "slit" experiment.
 |
 |
 |
It is necessary at this point to offer a bit of history about the development of the eye because the two-slit experiment represents a primitive model from which a primitive (and not necessarily pristine) view of the world can be seen. If this is the case, then the old two-slit experiment, having been modified to present a clearer image of reality by altering the sensitivity of the photographic plate (back wall on which images appear), thus enabling the initial observation of wave patterns to elucidate the existence of what appears to be individual electron particles. In other words, the mechanism by which physicists use to discern a clearer, more distinct image of atomic reality has been accomplished by evolving the mechanism of their visual ability to see. Again, the two-slit experiment and its modern improvements represent primitive developments seen in the evolution of the eye:
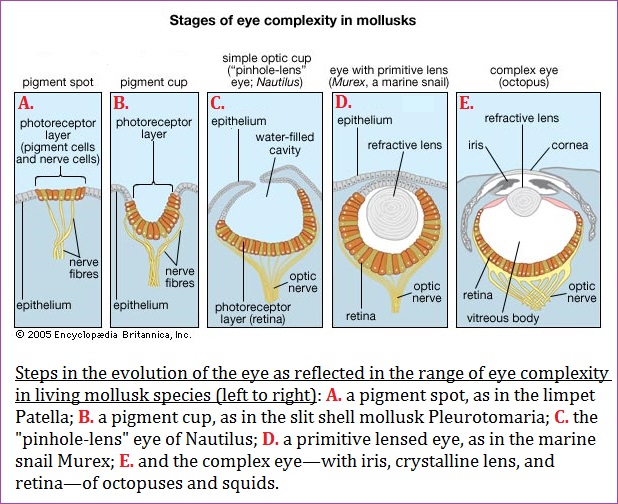
The soft-bodied animals that inhabited the world's seas before the Cambrian Period explosion (about 542 million years ago) undoubtedly had eyes, probably similar to the pigment-pit eyes of flatworms today. However, there is no fossil evidence to support the presence of eyes in the early soft-bodied creatures. Scientists know that the photo-pigment rhodopsin existed in the Cambrian Period. Evidence for this comes from modern metazoan phyla, which have genetically related rhodopsins, even though the groups themselves diverged from a common ancestor well before the Cambrian. 
By the end of the early Cambrian Period (roughly 521 million years ago), most, if not all, of the eye types in existence today had already evolved. The need for better eyesight arose because some of the animals in the early Cambrian fauna had turned from grazing to predation. Both predators and prey needed eyes to detect one another. Besides becoming better equipped visually, Cambrian animals developed faster forms of locomotion, and many acquired armoured exoskeletons, which have provided fossil material. Many of the animals in the famous Burgess Shale deposits in British Columbia, Can., had convex eyes that presumably had a compound structure. The best-preserved compound eyes from the Cambrian Period are found in the trilobites. Trilobite lenses were made of the mineral calcite, which enabled these organisms to fossilize exceptionally well. It is less certain when eyes of the camera-like single-chambered type first evolved. Fossil cephalopod mollusks appeared in the late Cambrian, and they probably had eyes resembling those of their present-day counterparts, such as lens eyes of Octopus or pinhole eyes of Nautilus. The first fish arose in the Ordovician Period (about 488 million to 444 million years ago) and radiated extensively in the Devonian Period (about 416 million to 359 million years ago). Fish fossils from these periods have eye sockets, indicating that these fish must have had eyes. The lampreys, present-day relatives of these early fish, have eyes that are very similar to those of other fish, leading to the conclusion that very little has happened to the aquatic form of the vertebrate eye for about 400 million years. The lower chordates, from which the vertebrates arose, have either simple eyespots or no eyes at all; therefore, presumably the vertebrate eye originated with the first fish and not before. Given the short time that eyes had to evolve in the Cambrian Period (some estimates of the explosive phase of the Cambrian radiation are as short as 20 million years), it is of some interest to know how long it would actually take an eye to evolve. British naturalist Charles Darwin was concerned about the difficulty of evolving an eye because it was “an organ of extreme perfection and complication.” Thus, it might be expected that eye evolution would take a long time. In 1994 Swedish zoologists Dan-Eric Nilsson and Susanne Pelger took up the challenge of “evolving” an eye of the fish type from a patch of photosensitive skin. Using pessimistic estimates of variation, heritability, and selection intensity, Nilsson and Pelger came to the conclusion that it would take 364,000 generations for a fish eye to evolve. Given a generation time of a year, which is typical for moderate-sized animals, a respectable eye could evolve in less than half a million years. Of course, other physiological elements (e.g., competent brains) have to evolve in parallel with eyes. However, at least as far as the eye itself is concerned, very little time is actually required for its evolution. Another problem concerning the evolution of eyes is the number of times eyes evolved. Given that the fossil record does not contain much information about the eyes of Precambrian animals, scientists have had to rely on evidence from the eyes of living Precambrian descendants to solve this problem. In 1977 Austrian zoologist Luitfried von Salvini-Plawen and American biologist Ernst Mayr examined the eyes and eyespots of representatives of all the main animal phyla and concluded that eyes of a basic kind had arisen independently at least 40 times and possibly as many as 65 times. The evidence presented by Salvini-Plawen and Mayr was of several kinds. At a cellular level, the receptive membrane of the photoreceptors could be elaborated from cilia or from microvilli (finger-like projections), the eyes could be derived either from epithelium or from nervous tissue, the axons of the receptors could leave from the back of the eye (everse) or from the front of the eye (inverse), and the overall eye design might be of the compound or the single-chambered type. Because these eye features tend to be stable within each phylum, the different combinations of features among phyla were taken to mean that the eyes had evolved independently. Set against this conclusion is the fact that some of the molecules involved in eye construction are indeed similar across phyla. The rhodopsin molecule itself is sufficiently similar among the vertebrates, the arthropods, and the cephalopod mollusks to rely on common ancestry as the most likely explanation for similarity in eye construction. A gene that is associated with eye development, Pax-6 (paired box gene 6), is very similar in insects and mammals, and it also occurs in the eyes of cephalopod mollusks. Thus, the earliest metazoans had at least some of the molecules necessary for producing eyes. These molecules were passed on to the metazoans' descendants, who used the molecules in different ways to produce eyes of very varying morphology. Because there are only a limited number of ways that images can be produced, it is not surprising that some of them have been “discovered” more than once. This has led to numerous examples of convergence in the evolutionary history of eyes. The similarity in optical design of the eyes of fish and cephalopod mollusks, such as octopuses and squid, is perhaps the most well-known example, but it is only one of many. The same lens design is also found in several groups of gastropod mollusks, in certain predatory worms (family Alciopidae), and in copepod crustaceans (genus Labidocera). A similar lens structure is also found in the extraordinary intracellular eye of a dinoflagellate protozoan (genus Warnowia). Compound eyes probably evolved independently in the chelicerata (genus Limulus), the trilobites, and the myriapods (genus Scutigera). Compound eyes appear to have evolved once or several times in the crustaceans and insects, in the bivalve mollusks (genus Arca), and in the annelid worms (genus Branchiomma). There are comparatively few cases in which one type of eye has evolved into a different type. However, it is thought that the single-chambered eyes of spiders and scorpions are descended from the compound eyes of earlier chelicerates (e.g., genus Limulus, eurypterids) by a process of reduction. Something similar has occurred in the amphipod crustacean genus Ampelisca, where single-chambered eyes have replaced the compound eyes typical of the group. Michael Land: Emeritus professor of neurobiology, Sussex Vision Laboratory, Sussex Centre for Neuroscience, University of Sussex, Brighton, England. Source: "Photoreception." Encyclopædia Britannica Ultimate Reference Suite, 2013. |
If we compare the "visual apparatus" used in the two-split experiment and that of a human eye, the differences in complexity are not difficult to point out... except that we should point out that if simplicity was the rule of thumb for evolutionary processes, there would be no need for complexity in cellular structure or species' development. If we were to replicate the complexity (presumed simplicity) of the two human eyes into the two-slit experiment (taking into consideration the concept of the "third eye" denoted as the pineal gland); a different atomic reality might well unfold for us to see... if we permit ourselves to get past our primitive means of visualizing it. In other words, and even though some would argue the necessity of simplifying experiments, equations, etc., such a perspective may be due to internalized inclinations that remain attached to a primitive past of conceptualization. We use a simple experiment to arrive at a simplistic explanation of relativity because the brains of physicists are most comfortable seeing the world through a primitive lens of appreciation to which they label with other (numerical) symbolism like the divination ceremonies of old which entertained the reality of discerning reality by investigating tea leaves, tarot cards, entrails, ashes, clouds, insect movement, bird flights, etc..., assigned with their own respective symbolism.
 |
 |
Yet, let us ask whether the world of atomic particles is a world of light and dark bands, or does it have color? Do the names used for some particles (strange, up/down, etc...) actually reflect the reality of the atomic world or is it more of the same human primitiveness being used? If there is color, if there actually is strangeness and direction, should not our experiments reflect the reality of the atomic world and not necessarily the world humans are or aren't capable of seeing correctly? And what of the spacing of the slits? Is there a correlation with biological arrangements of eyes that can or even should be made? Let's look at a short Britannica excerpt about eye types and placement:
|
The eyes of animals are diverse not only in size and shape but also in the ways in which they function. For example, the eyes of fish from the deep sea often show variations on the basic spherical design of the eye. In these fish, the eye's field of view is restricted to the upward direction, presumably because this is the only direction from which there is any light from the surface. This makes the eye tubular in shape. Some fish living in the deep sea have reduced eyelike structures directed downward (e.g., Bathylychnops, which has a second lens and retina attached to the main eye); it is thought that the function of these structures is to detect bioluminescent creatures. On the ocean floor, where no light from the sky penetrates, eyes are often reduced or absent. However, in the case of Ipnops, which appears to be eyeless, the retina is still present as a pair of plates covering the front of the top of the head, although there is no lens or any other optical structure. The function of this eye is unknown. 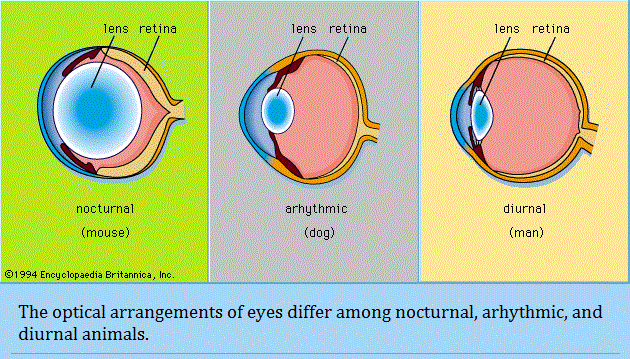 
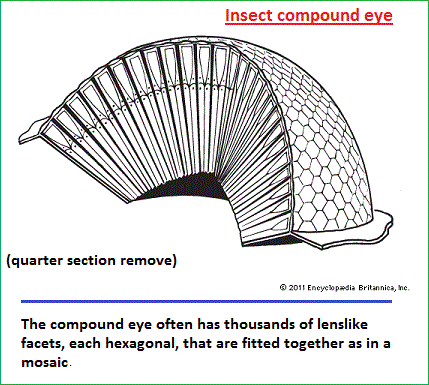
The placing of the eyes in the head varies. Predators, such as felines and owls, have forward-pointing eyes and the ability to judge distance by binocular triangulation. Herbivorous species that are likely to be victims of predation, such as mice and rabbits, usually have their eyes almost opposite each other, giving near-complete coverage of their surroundings. In addition to placement in the head, the structure of the eye varies among animals. Nocturnal animals, such as the house mouse and opossum, have almost spherical lenses filling most of the eye cavity. This design allows the eye to capture the maximum amount of light possible. In contrast, diurnal animals, such as humans and most birds, have smaller, thinner lenses placed well forward in the eye. Nocturnal animals usually have retinas with a preponderance of photoreceptors called rods, which do not detect colour but perceive size, shape, and brightness. Strictly diurnal animals, such as squirrels and many birds, have retinas containing photoreceptors called cones, which perceive both colour and fine detail. A slit pupil is common in nocturnal animals, as it can be closed more effectively in bright light than a round pupil. In addition, nocturnal animals, such as cats and bush babies, are usually equipped with a tapetum lucidum, a reflector behind the retina designed to give receptors a second chance to catch photons that were missed on their first passage through the retina. Animals such as seals, otters, and diving birds, which move from air to water and back, have evolved uniquely shaped corneas—the transparent membrane in front of the eye that separates fluids inside the eye from fluids outside the eye. The cornea functions to increase the focusing power of the eye; however, optical power is greatly reduced when there is fluid on both sides of the membrane. As a result, seals, which have a nearly flat cornea with little optical power in air or water, rely on a re-evolved spherical lens to produce images. Diving ducks, on the other hand, compensate for the loss of optical power in water by squeezing the lens into the bony ring around the iris, forming a high curvature blip on the lens surface, which shortens its focal length (the distance from the retina to the centre of the lens). One of the most interesting examples of amphibious optics occurs in the “four-eyed fish” of the genus Anableps, which cruises the surface meniscus with the upper part of the eye looking into air and the lower part looking into water. It makes use of an elliptical lens, with the relatively flat sides adding little to the power of the cornea and the higher curvature ends focusing light from below the surface, where the cornea is ineffective. Though the eyes of animals are diverse in structure and use distinct optical mechanisms to achieve resolution, eyes can be differentiated into two primary types: single-chambered and compound. Single-chambered eyes (sometimes called camera eyes) are concave structures in which the photoreceptors are supplied with light that enters the eye through a single lens. In contrast, compound eyes are convex structures in which the photoreceptors are supplied with light that enters the eye through multiple lenses. The possession of multiple lenses is what gives these eyes their characteristic faceted appearance. Michael Land: Emeritus professor of neurobiology, Sussex Vision Laboratory, Sussex Centre for Neuroscience, University of Sussex, Brighton, England.Source: "Photoreception." Encyclopædia Britannica Ultimate Reference Suite, 2013. |
The above excerpt doesn't really do justice to the volume and value of the whole article by Michael Land. For those interested in constructing a better experiment than the double-slit model, it should be consulted as an introduction to an appreciation of the eye and vision. For example, how many reflectors for receptors do we humans use in the double-slit experiments that might be better if they were altered to represent the evolutionary stages of the eye... and into the future as resources dwindle, and the surface of Earth dramatically changes, thereby calling for alterations in visual perception? No less, is a visual oriented model the proper route for getting a better grasp of atomic reality? If we use a non-visual recording devise, will there be something lost in the translation between it and an attempt to render a visual image? In other words, if it is more appropriate to use a "music of the spheres" means of identifying the atomic world, will an attempt to translate this into a visual illustration reproduce a crude and unreliable picture?
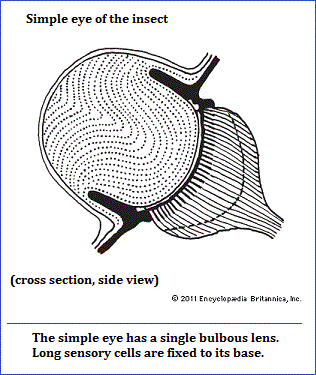
Is a "forward facing" two-slit arrangement the best way to see the atomic world? Is a slit, a round pupil, or some other configuration a better (aperture)? Should we use multiple forms or one in a series? Are we seeing particles and waves or is this merely the adopted way we chosen to describe the images according to the type of experimental configuration we use? Are the assumed waves and particles the result of the type of medium we're using because we still think in terms of primates trying to fish out insects from a hole with a twig, that other primates want to describe as tool usage? Why has biology been limited to the usage of a small range of light receiving forms, when there are many other models to choose from?... as seen in the following image:

Why has the line (slit), circle (circular eyeball), and triangular-like binocular vision arrangements been used amongst all the many different types of shapes? Whereas one may think in terms of biological constraints, or some childish theological reason, the point to be made is that there is a conservation. And this conservation may have nothing to do with the reality of atomic existence, but is a requirement of an adaptive species to adopt the best means of survival in an environment headed for decay. As such, all our explanations for the reality of atomic phenomena are part of the rationalized adjustments we use to make the best of an increasingly bad situation... namely, the incremental decay of the planet.
On the one hand we try to unravel the world of atomic phenomena, and yet resort to the usage of a primitive type of perception that appears remarkably like that of a simple eye. And even though the above comments by Michael Land denote two primary types of eyes labeled simple and compound, this reference also refers to the oft'- used "1-—many" assemblage of compartmentalization found in other subject areas, and presents us with another instance for the existence of a body of conservation related to an increasing usage of conservative perspectives in an attempt to deal with diminishing resources. In other words, while we're trying to uncover the fundamental laws of nuclear particles, we are being forced to carry out the experiments and accompanying ideological rules-of-thumb in an environment that is forcing us into a conservative mode of operation. Such an environment may not be what is best for uncovering that which we seek to know. It's like trying to write down knowledge contained on a rockface while having to dodge the effects of an ongoing volcano and earthquake. The double-slit experiment and the application of what has been found is like trying to catch a hot potato that's getting hotter, yet some physicists think they have constructed a suitable pair of gloves because it's part of an adopted rational because no one has come up with anything better that they want to agree with.
The eyes of biological organisms are not static... are not still. In other words, if they don't move about by muscles, reflexive actions, etc., then they are attached to an organism, a life form that is mobile. In a double-slit experiment, the eyes are fixed. the light source is fixed and so is the receptor screen, plate or wall. In terms of human perception, an illuminated object in mind does not become the object before one's eyes in a similar manner. In other words, we haven't as yet been able to reverse the process. The same goes for the double-slit experiment. Whereas we may see particles at one end having been derived from waves coming from a light source, the process is not customarily reversed. We don't take a light source, screen it through two slits that are then subjected to a further screening process to yield a particle from which we derive the concept of waves and particles as being two separate ingredients. Of course there might very well be two entities if our minds are predisposed to seeing the world in groups of two... as part of an anciently established conceptualization of the world to which the concept of the Peace/War dichotomy obviously belong.
Interestingly, if you look at the slit of the sheep in the following image, you may not only make note of the slit, but that the slit remains in the same relative (horizontal) position to the ground. The image next to it shows two different reptile eyes. If it is not because of some conservation effect, why can't the pupil be in the shape of another form? For example, it might be appropriate for those who lust after money to have dollar signs for pupils, or those in love to have heart-shaped pupils, book readers to have rectangular eyes, etc...
 |
 |
 | |
 |
 |
We'll continue with the present topic of the double-split experiment and how it relates to the dichotomy of the Peace/War model of conceptualization on the next page.
Date of Origination: Friday, 20-Jan-2017... 04:20 AM
Date of initial posting: Saturday, 28-Jan-2017... 12:54 PM
Updated Posting: Monday, 12-June-2017... 4:24 AM
Updated posting: Friday, 6-April-2018... 12:27 PM