page 28
Note: the contents of this page as well as those which precede and follow, must be read as a continuation and/or overlap in order that the continuity about a relationship to/with the dichotomous arrangement of the idea that one could possibly talk seriously about peace from a different perspective as well as the typical dichotomous assignment of Artificial Intelligence (such as the usage of zeros and ones used in computer programming) ... will not be lost (such as war being frequently used to describe an absence of peace and vice-versa). However, if your mind is prone to being distracted by timed or untimed commercialization (such as that seen in various types of American-based television, radio, news media and magazine publishing... not to mention the average classroom which carries over into the everyday workplace), you may be unable to sustain prolonged exposures to divergent ideas about a singular topic without becoming confused, unless the information is provided in a very simplistic manner.
Let's face it, humanity has a lousy definition, accompanying practice, and analysis of peace.
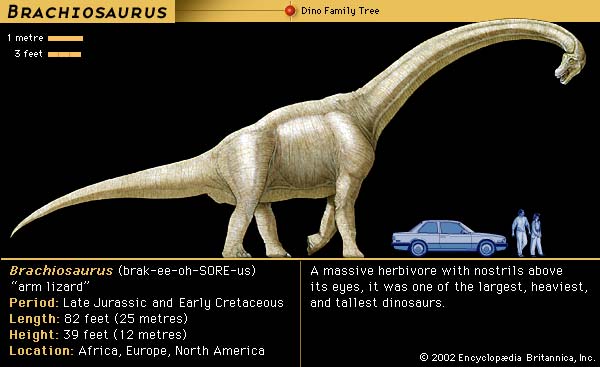
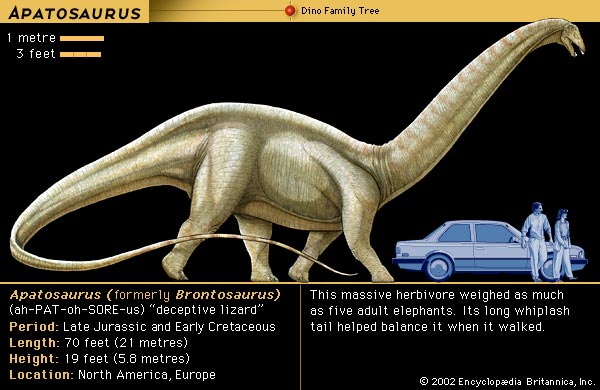
If we look upon the Peace/War (conflict) duality as a generalized theory instead of as a featured law of life or natural event on this planet, and then align the two with the words Wave and Particle, what we have is a reverse-engineered diagram of an experimental apparatus that hasn't been identified because we are too close to it... like an ant having climbed several hills only to find out later, while at a greater distance away, that we had stood on the head of a Brachiosaurus or Brontosaurus, (later changed to Apatosaurus).
|
 |
In presenting other examples of dinosaurs in the present context, we can view them as experiments carried out over long expanses of time (with respect to a human life scale). For whatever reason(s), primates became one of the main experiments which continues to this day. And though we don't think of the existence of Existence as an experiment, it too might well be... without having to fall into the bog of associating the word "experiment" with an "experimenter", since there may be more than one, only one, or merely refers to a process from which lowly primate creatures such as humans like to infer sentient causality. No less, nuclear forces and the electromagnetic spectrum also may be experiments to which are currently experiencing but one model thereof. They could be experiments which are part of a series of events which come and go, like the cyclical nature of seasons, to which the concepts peace and war may belong to the same pendulum.
The rationale we use for defining an alignment of primates may not work for us, even in the most general terms, in the same manner if applied to non-primate species... much less (seemingly) inanimate circumstances, but allows us to stretch the malleability of our minds, so to speak, enabling other wayward thoughts to appear serendipitously if such a soil is prepared. Creative thinking often requires cultivation of alternative perspectives that we don't actually have to believe in, but allow the contours of musefulness to "play in the sand" and then bring it into a more conventional box-like conceptualization of testability. In such a way, we permit ourselves to explore alternative perspectives of the Peace/War dichotomy by taking a step back from it through a formula of mixing and matching, deletion, substitution, and the characteristic basic math operations (+, /, X, -).
Words such as convergence, divergence, repetition, series, parallel, correspondence, comparison, etc., are as much mathematical terms as they are in describing events in literary tales. The problem which frequently arises in developing new ideas is not that there are ample sources for getting different ideas, but that internalized (one's self, one's friends, one's classmates, one's colleagues, one's family, one's relatives, one's neighbors, one's culture, one's religion, one's workplace, etc...) or externalized (academic genre, traditions, dictionary outlined definitions, etc...) restrictions may be imposed. For example, comparing insect and human behavior may be frowned on by someone in your respective social circle, or claiming that a tomato and chili are fruits instead of vegetables may be derided by one's parents, relatives and their friends. The same, or similar experiment (or at least basic general idea) used in one area of application might well be applied with different ingredients such as when baking or cooking. In a sense, one cooks up a new idea using an old method (pan or pot) with different ingredients... adjusting the flame of the burner by way of a measured or happenstance methodology.
The same type of experimentation or similar experimental process can take place in different places in different time periods though circumstances appear to be different. For example, experiments in genetic hybridization occurred amongst several researchers, but Gregor Mendel is credited with being the first to discover certain key characteristics, even though those whom he relayed the information to did not grasp the importance of what he had found. Others did not have the correct frame of mind for understanding the meaning of his research. There are of course other examples, but let us remain on the trek of evolution for a bit by providing an excerpt from the Britannica on topics explaining the processes of evolution from different perspectives (as if one were witnessing not different ideas but different brain functionings related to evolutionary changes within, between and in spite of socialization in different or similar environments).[ Inserted notes and commentary from this writer in the Peace series are in blue:]
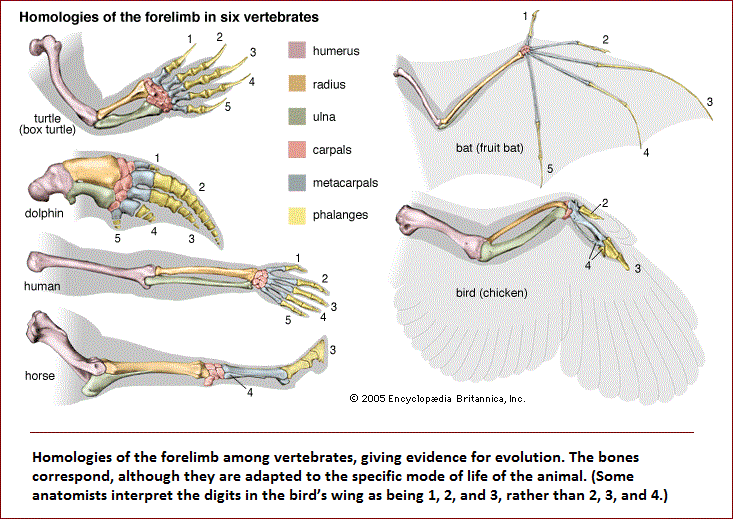
| Convergent and parallel evolution A distinction has to be made between resemblances due to propinquity (being close together) of descent and those due only to similarity of function. Structural similarities, correspondence of features in different organisms that is due to inheritance from a common ancestor is called homology. The fore-limbs of humans, whales, dogs, and bats are homologous. The skeletons of these limbs are all constructed of bones arranged according to the same pattern because they derive from a common ancestor with similarly arranged fore-limbs. Correspondence of features due to similarity of function but not related to common descent is termed analogy. The wings of birds and of flies are analogous. Their wings are not modified versions of a structure present in a common ancestor but rather have developed independently as adaptations to a common function, flying. The similarities between the wings of bats and birds are partially homologous and partially analogous. Their skeletal structure is homologous, due to common descent from the fore-limb of a reptilian ancestor; but the modifications for flying are different and independently evolved, and in this respect they are analogous. |
Yet, why didn't all life forms develop wings? Why have different life forms at all? Why have different electromagnetic radiation levels? Is the wave/particle form and functionality a generalist way of survival, much like humans that have survived by adopting multiple traits, characters, behaviors, talents, etc.? Why have different atomic particles? Because of the type of ancestry? Where as the old model of humanity was divided into three races (African- Asian- Caucasian), and there are three families of fundamental particles (stable, unstable, highly unstable) and three fundamental particles (electrons- neutrons- protons), etc., are there further correlations of relevance to make such that one race is more or less stable or highly unstable than the rest?a Does this apply to different forms of radiation as well, or are they divergent characteristics of the same species of radiation that has a common ancestor? Do all atomic particles have a common ancestry and their present form is due to the type of environment its ancestry was subjected to? Can such (inanimate) species be changed by altering their environment? Can we do the same for the Peace/War dichotomy which may not be a dichotomy but simply different perspectives of the same species of "elephant"?(See previous page in this series for a reference to the word "elephant" being used as a metaphor.) |
 Features that become more rather than less similar through independent evolution are said to be convergent. Convergence is often associated with similarity of function, as in the evolution of wings in birds, bats, and flies. The shark (a fish) and the dolphin (a mammal) are much alike in external morphology; their similarities are due to convergence, since they have evolved independently as adaptations to aquatic life. Taxonomists also speak of parallel evolution. Parallelism and convergence are not always clearly distinguishable. Strictly speaking, convergent evolution occurs when descendants resemble each other more than their ancestors did with respect to some feature. Parallel evolution implies that two or more lineages have changed in similar ways, so that the evolved descendants are as similar to each other as their ancestors were. The evolution of marsupials in Australia, for example, paralleled the evolution of placental mammals in other parts of the world. There are Australian marsupials resembling true wolves, cats, mice, squirrels, moles, groundhogs, and anteaters. These placental mammals and the corresponding Australian marsupials evolved independently but in parallel lines by reason of their adaptation to similar ways of life. Some resemblances between a true anteater (genus Myrmecophaga) and a marsupial anteater, or numbat (Myrmecobius), are due to homology—both are mammals. Others are due to analogy—both feed on ants. |
Let us also look at this from an architectural perspective. For example, it was found that ancient Egyptians used smaller models of pyramids that we of today interpret to have been construction aids for larger efforts... and appear as small stepped pyramids— a design used by others as shown in the two accompanying images:
 |
 |
 |
 |
Let us note that children's toys often reference the life-size (and sometimes animate) models in a reverse order of development (life-sized models used as illustrations for the designing of toys); the development of a particular toy design did not necessarily precede the development of the life-size model; although the stacking of blocks (or other available objects in a given quantity) appears to be a rather universal characteristic amongst children... and the later usage of stacking blocks to build pyramids may be little more than an adult expression of a childhood activity. No less, it should be entered into the account that archaeologists have (it is said), uncovered toys with wheels attached but have not found such an idea adapted to usage by adults in their everyday routines. Though such life-sized wheel craft may have been used, there are no representations of life-size models... not with the invention of an axle anyway. Analogously, evolution can have small versions of a given organism that do not become a full-sized model and full-sized models may not "evolve" into smaller versions. For example, while pterodactyls were large, they were not appreciably so in comparison to other four-legged behemoths, yet we of the present day can identify very small bird versions such as the humming bird. And though one might want to make a case for the presence of large dinosaurs in the form of large construction equipment, we would then have to consider that evolution of biology also includes a possible lineage of inanimate forms.
 |
 Fred Flintstone and Barney Rubble may work here |
|
Parallel and convergent evolution are also common in plants. New World cacti and African euphorbias, or spurges, are alike in overall appearance although they belong to separate families. Both are succulent, spiny, water-storing plants adapted to the arid conditions of the desert. Their corresponding morphologies have evolved independently in response to similar environmental challenges. 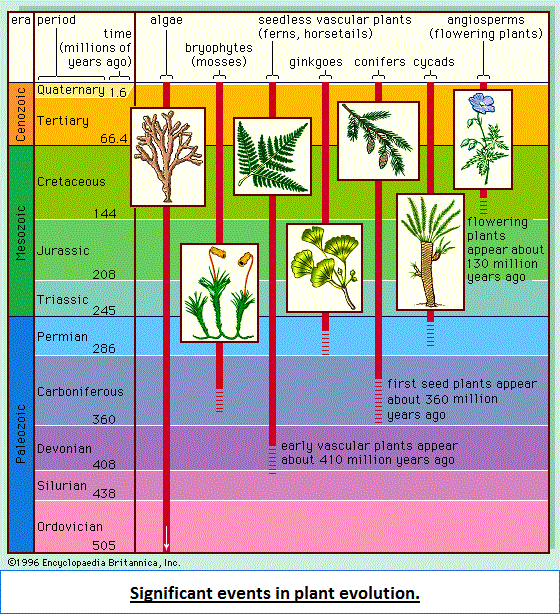 Homology can be recognized not only between different organisms but also between repetitive structures of the same organism. This has been called serial homology. There is serial homology, for example, between the arms and legs of humans, between the seven cervical vertebrae of mammals, and between the branches or leaves of a tree. The jointed appendages of arthropods are elaborate examples of serial homology. Crayfish have 19 pairs of appendages, all built according to the same basic pattern but serving diverse functions—sensing, chewing, food handling, walking, mating, egg carrying, and swimming. Although serial homologies are not useful in reconstructing the phylogenetic relationships of organisms, they are an important dimension of the evolutionary process. | ||
| ||
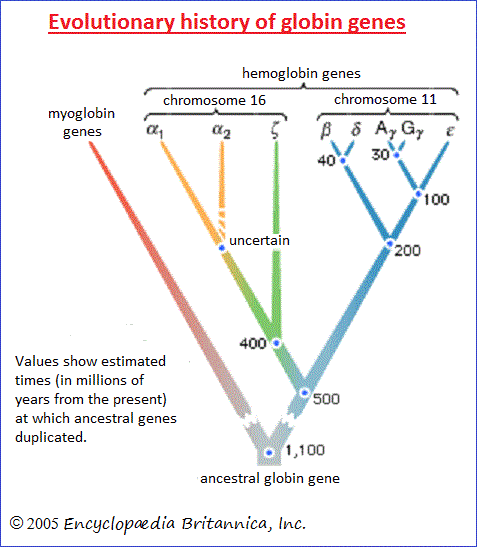
Relationships in some sense akin to those between serial homologs exist at the molecular level between genes and proteins derived from ancestral gene duplications. The genes coding for the various hemoglobin chains are an example. About 500 million years ago a chromosome segment carrying the gene coding for hemoglobin became duplicated, so that the genes in the different segments thereafter evolved in somewhat different ways, one eventually giving rise to the modern gene coding for the α hemoglobin chain, the other for the β chain. The β chain gene became duplicated again about 200 million years ago, giving rise to the γ hemoglobin chain, a normal component of fetal hemoglobin (hemoblobin F). The genes for the α, β, γ, and other hemoglobin chains are homologous; similarities in their nucleotide sequences occur because they are modified descendants of a single ancestral sequence. There are two ways of comparing homology between hemoglobins. One is to compare the same hemoglobin chain—for instance, the α chain—in different species of animals. The degree of divergence between the α chains reflects the degree of the evolutionary relationship between the organisms, because the hemoglobin chains have evolved independently of one another since the time of divergence of the lineages leading to the present-day organisms. A second way is to make comparisons between, say, the α and β chains of a single species. The degree of divergence between the different globin chains reflects the degree of relationship between the genes coding for them. The different globins have evolved independently of each other since the time of duplication of their ancestral genes. Comparisons between homologous genes or proteins within a given organism provide information about the phylogenetic history of the genes and hence about the historical sequence of the gene duplication events. Whether similar features in different organisms are homologous or analogous—or simply accidental—cannot always be decided unambiguously, but the distinction must be made in order to determine phylogenetic relationships. Moreover, the degrees of homology must be quantified in some way so as to determine the propinquity of common descent between species. Difficulties arise here as well. In the case of forelimbs, it is not clear whether the homologies are greater between human and bird than between human and reptile, or between human and reptile than between human and bat. The fossil record sometimes provides the appropriate information, even though the record is deficient. Fossil evidence must be examined together with the evidence from comparative studies of living forms and with the quantitative estimates provided by comparative studies of proteins and nucleic acids. |


The same goes for when making a comparison between sports such as football and soccer, flying machines, fishing poles, hunting/herding/butchering techniques, definitions of right, wrong, good, bad, peace, war— or when examining religions... such as in the case of a specific attribute regarding an anthropomorphic representation of evil, angels, or a god... though morality may one day be personified into a human form as well.
Gradual and punctuational evolution
The fossil record indicates that morphological evolution is by and large a gradual process. Major evolutionary changes are usually due to a building-up over the ages of relatively small changes. But the fossil record is discontinuous. Fossil strata are separated by sharp boundaries; accumulation of fossils within a geologic deposit (stratum) is fairly constant over time, but the transition from one stratum to another may involve gaps of tens of thousands of years. Whereas the fossils within a stratum exhibit little morphological variation, new species—characterized by small but discontinuous morphological changes—typically appear at the boundaries between strata. That is not to say that the transition from one stratum to another always involves sudden changes in morphology; on the contrary, fossil forms often persist virtually unchanged through several geologic strata, each representing millions of years. The apparent morphological discontinuities of the fossil record are often attributed by paleontologists to the discontinuity of the sediments— that is, to the substantial time gaps encompassed in the boundaries between strata. The assumption is that, if the fossil deposits were more continuous, they would show a more gradual transition of form. Even so, morphological evolution would not always keep progressing gradually, because some forms, at least, remain unchanged for extremely long times. Examples are the lineages known as "living fossils"—for instance, the lamp shell Lingula, a genus of brachiopod (a phylum of shelled invertebrates) that appears to have remained essentially unchanged since the Ordovician Period, some 450 million years ago; or the tuatara (Sphenodon punctatus), a reptile that has shown little morphological evolution for nearly 200 million years, since the early Mesozoic. Some paleontologists have proposed that the discontinuities of the fossil record are not artifacts created by gaps in the record but rather reflect the true nature of morphological evolution, which happens in sudden bursts associated with the formation of new species. The lack of morphological evolution, or stasis, of lineages such as Lingula and Sphenodon is in turn due to lack of speciation within those lineages. The proposition that morphological evolution is jerky, with most morphological change occurring during the brief speciation events and virtually no change during the subsequent existence of the species, is known as the punctuated equilibrium model. Whether morphological evolution in the fossil record is predominantly punctuational or gradual is a much-debated question. The imperfection of the record makes it unlikely that the issue will be settled in the foreseeable future. Intensive study of a favourable and abundant set of fossils may be expected to substantiate punctuated or gradual evolution in particular cases. But the argument is not about whether only one or the other pattern ever occurs; it is about their relative frequency. Some paleontologists argue that morphological evolution is in most cases gradual and only rarely jerky, whereas others think the opposite is true. Much of the problem is that gradualness or jerkiness is in the eye of the beholder. Consider the evolution of shell rib strength (the ratio of rib height to rib width) within a lineage of fossil brachiopods of the genus Eocelia. Results of the analysis of an abundant sample of fossils in Wales from near the beginning of the Devonian Period is shown in the figure. One possible interpretation of the data is that rib strength changed little or not at all from 415 million to 413 million years ago; rapid change ensued for the next 1 million years, followed by virtual stasis from 412 million to 407 million years ago; and then another short burst of change occurred about 406 million years ago, followed by a final period of stasis. On the other hand, the same record may be interpreted as not particularly punctuated but rather a gradual process, with the rate of change somewhat greater at particular times. 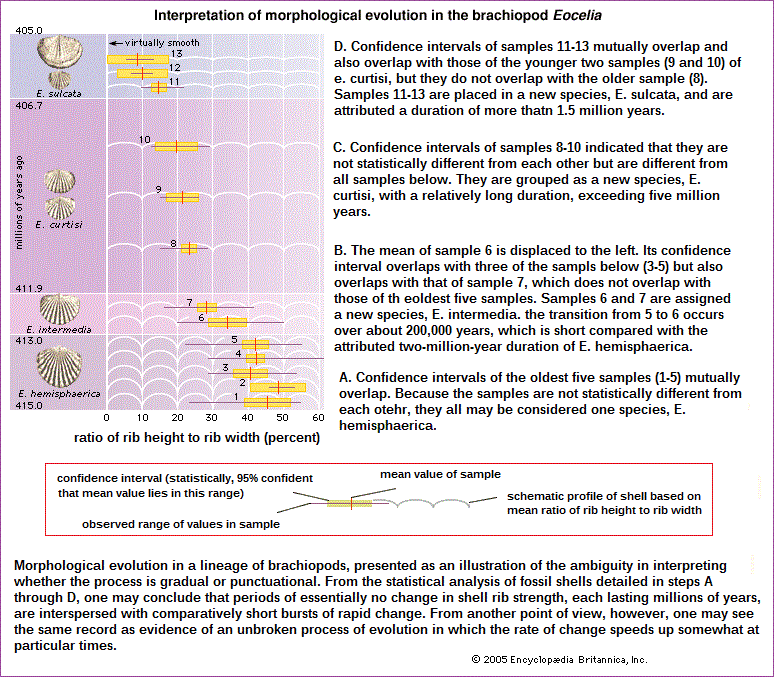 The proponents of the punctuated equilibrium model propose not only that morphological evolution is jerky but also that it is associated with speciation events. They argue that phyletic evolution—that is, evolution along lineages of descent—proceeds at two levels. First, there is continuous change through time within a population. This consists largely of gene substitutions prompted by natural selection, mutation, genetic drift, and other genetic processes that operate at the level of the individual organism. The punctualists maintain that this continuous evolution within established lineages rarely, if ever, yields substantial morphological changes in species. Second, they say, there is the process of origination and extinction of species, in which most morphological change occurs. According to the punctualist model, evolutionary trends result from the patterns of origination and extinction of species rather than from evolution within established lineages.  Speciation involves the development of reproductive isolation between populations previously able to interbreed. Paleontologists discriminate between species by their different morphologies as preserved in the fossil record, but fossils cannot provide evidence of the development of reproductive isolation—new species that are reproductively isolated from their ancestors are often morphologically indistinguishable from them. Speciation as it is seen by paleontologists always involves substantial morphological change. This situation creates an insuperable difficulty for resolving the question of whether morphological evolution is always associated with speciation events. If speciation is defined as the evolution of reproductive isolation, the fossil record provides no evidence that an association between speciation and morphological change is necessary. But if new species are identified in the fossil record by morphological changes, then all such changes will occur concomitantly with the origination of new species. 
For those readers who do not have difficulty in "translating" the basic ideas from one subject to another in order to make comparisons, we can interject the topic of languages because the word "family" has frequently been used to describe different languages grouped according to one or another characteristic such as the Afro-Asiatic, Tai, Romance, Native American, Chinese, etc... languages. And though we don't think of particular ideas representing an independent language, it might be helpful in our peace/war research topic since it might yield a fruitful appreciation that what we are dealing with are different languages of the mind/brain, related to values which can be enumerated, such as mono, di/bi, tri... etc... In other words, what if a perspective involving dichotomies involved understanding the formula as if it were a language that is similar to, but appreciably different than trichotomies, singularities, etc...? With such a perspective in mind, the biblical idea about a Tower of Babel arises not because languages or speech is unintelligible, but because we are unable to accurately distinguish what is and is not a language... for example, the expressions or utterances of mathematics, music, art, logic, and other non-typically defined "languages". Indeed, many a parent has stressed over not being able to understand the language of the "new math" their first grader has brought home as homework. No less, the usage of the metric system has been treated as a foreign language by many American... not to mention modern art, rap and other creatively expressed perspectives. And we could include the "language" of graffiti, gang signs, and the nonsense so many politicians have a knack for professing as some truth. Not to mention the disconcertions of many a man trying to interpret what a woman is actually saying, particularly in those instances if she herself doesn't quite know yet even when something is said... due to not wanting to make a commitment in order to provide herself an opportunity to take advantage of that which may or may not present itself... she will at least have the option to say (with her eyes or quirky smile) that she told you so. |
|
Diversity and extinction The current diversity of life is the balance between the species that have arisen through time and those that have become extinct. Paleontologists observe that organisms have continuously changed since the Cambrian Period, more than 500 million years ago, from which abundant animal fossil remains are known. The division of geologic history into a succession of eras and periods (see figure) is hall-marked by major changes in plant and animal life—the appearance of new sorts of organisms and the extinction of others. Paleontologists distinguish between background extinction, the steady rate at which species disappear through geologic time, and mass extinctions, the episodic events in which large numbers of species become extinct over time spans short enough to appear almost instantaneous on the geologic scale. 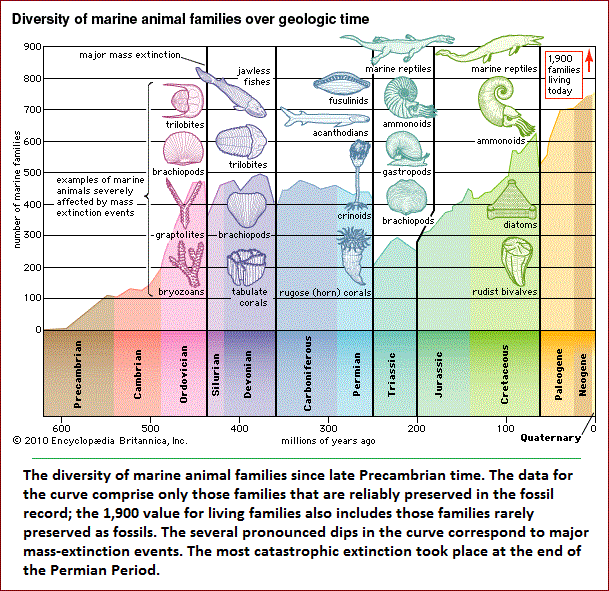 Best known among mass extinctions is the one that occurred at the end of the Cretaceous Period, when the dinosaurs and many other marine and land animals disappeared. Most scientists believe that the Cretaceous mass extinction was provoked by the impact of an asteroid or comet on the tip of the Yucatán Peninsula in southeastern Mexico 65 million years ago. The object's impact caused an enormous dust cloud, which greatly reduced the Sun's radiation reaching Earth, with a consequent drastic drop in temperature and other adverse conditions. Among animals, about 76 percent of species, 47 percent of genera, and 16 percent of families became extinct. Although the dinosaurs vanished, turtles, snakes, lizards, crocodiles, and other reptiles, as well as some mammals and birds, survived. Mammals that lived prior to the event were small and mostly nocturnal, but during the ensuing Paleogene and Neogene periods they experienced an explosive diversification in size and morphology, occupying ecological niches vacated by the dinosaurs. Most of the orders and families of mammals now in existence originated in the first 10 million—20 million years after the dinosaurs' extinction. Birds also greatly diversified at that time. Several other mass extinctions have occurred since the Cambrian. The most catastrophic happened at the end of the Permian Period, about 251 million years ago, when 95 percent of marine species, 82 percent of genera, and 51 percent of families of animals became extinct. (See also Triassic Period: Permian-Triassic extinctions.) Other large mass extinctions occurred at or near the end of the Ordovician (about 444 million years ago, 85 percent of marine species extinct), Devonian (about 359 million years ago, 70-80 percent of species extinct), and Triassic (about 200 million years ago, nearly 80 percent of species extinct). Changes of climate and chemical composition of the atmosphere appear to have caused these mass extinctions; there is no convincing evidence that they resulted from cosmic impacts. Like other mass extinctions, they were followed by the origin or rapid diversification of various kinds of organisms. The first mammals and dinosaurs appeared after the late Permian extinction, and the first vascular plants after the Late Ordovician extinction. Background extinctions result from ordinary biological processes, such as competition between species, predation, and parasitism. When two species compete for very similar resources—say, the same kinds of seeds or fruits—one may become extinct, although often they will displace one another by dividing the territory or by specializing in slightly different foods, such as seeds of a different size or kind. Ordinary physical and climatic changes also account for background extinctions—for example, when a lake dries out or a mountain range rises or erodes. New species come about by the processes discussed in previous sections. These processes are largely gradual, yet the history of life shows major transitions in which one kind of organism becomes a very different kind. The earliest organisms were prokaryotes, or bacteria-like cells, whose hereditary material is not segregated into a nucleus. Eukaryotes have their DNA organized into chromosomes that are membrane-bound in the nucleus, have other organelles inside their cells, and reproduce sexually. Eventually, eukaryotic multicellular organisms appeared, in which there is a division of function among cells—some specializing in reproduction, others becoming leaves, trunks, and roots in plants or different organs and tissues such as muscle, nerve, and bone in animals. Social organization of individuals in a population is another way of achieving functional division, which may be quite fixed, as in ants and bees, or more flexible, as in cattle herds or primate groups. Because of the gradualness of evolution, immediate descendants differ little, and then mostly quantitatively, from their ancestors. But gradual evolution may amount to large differences over time. The fore-limbs of mammals are normally adapted for walking, but they are adapted for shoveling earth in moles and other mammals that live mostly underground, for climbing and grasping in arboreal monkeys and apes, for swimming in dolphins and whales, and for flying in bats. The fore-limbs of reptiles became wings in their bird descendants. Feathers appear to have served first for regulating temperature but eventually were co-opted for flying and became incorporated into wings. Francisco Jose Ayala: Donald Bren Professor of Biological Sciences, University of California, Irvine. Author of Evolving: The Theory and Processes of Organic Evolution and others.Source: "Evolution." Encyclopædia Britannica Ultimate Reference Suite, 2013. |
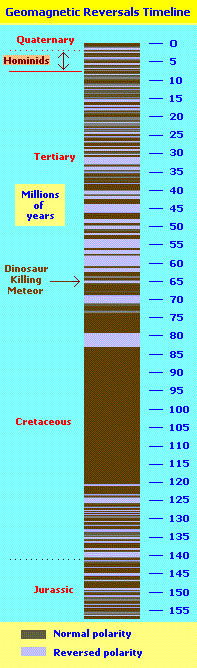
In illustrations describing the development of life or major extinction events (that have been uncovered), what is missing from these types of images is a record of Earth's magnetic reversals and the rate at which the Earth was revolving. Far too often we find that such depictions are interested in displaying the bulk of life forms from about 600 million years ago to the present. However, it should be noted that at our present stage of trying to get a firm grasp on the rotation rate game, our interpretations are much like trying to predict the weather, or some weather condition. Whereas a geologist, vulcanologist, or meteorologist can get it right sometimes, they don't get it right all the time. Here is a list of the values used in trying to determine the rate at which the Earth's rotation is slowing (and thus can be used to determine how fast the Earth was spinning at its formation. A "fluctuating" system of values were used in order to bring estimates in alignment with other evidence such as counting the growth layers of corals from the Devonian era. (The following short list provides different values that were found when researching data on how fast the Earth's rotation is slowing, in order to work out some semblance of a calculation to encompass the history of Earth. The table following the list represents different values of what this might look like over the history of the Earth... taking into account the need for making "adjustments" due to events which change the numbers. And in other words means that the slowing may not be constant over long stretches of time.
- 0.000015 seconds per day per year at present,
although there is reason to think that over long periods in the past the slightly
higher figure of 0.000020 is more accurate. 1/100 sec/century, because of tidal
effects!
- 0.000020 seconds per day per year in the past
- .00002 seconds per day per year
- .00002 seconds per year
- .0002 seconds per day per year
- .0015 seconds every second
- .0016 seconds per century (1.6 milliseconds)
- .0017 msec per day per century (1.7 milliseconds)
- .001 seconds per century
- .002 seconds per day
- .002 seconds per day per year
- .002 seconds per century (2 milliseconds)
- .005 seconds per century (5 milliseconds)
- .0053 seconds per year
- 1.4 milliseconds per century
- 1.48 milliseconds per century
- 1.5 milliseconds per day (0.0015)
- 1.5 milliseconds a day per day (some creationists view this as correct)
- 1.5 milliseconds a day per century
- 1.5 milliseconds per century
- 1.5 - 2 milliseconds per century
- 1.7 milliseconds per year
- 2.3 milliseconds per century
- 2.5 milliseconds per century
- 3 milliseconds per century
- 1 second every 500 days
- 1 second every ten years
- 1 second every 625 years
- 1 second every 50,000 years
- 2.2 seconds per 100,000 years
- 16 seconds every million years
- 22 seconds every million years
- 30 seconds per century
- 3 hours over 2000 years
- 20 days per year every 200 million years
- etc...
Using a small hand-held calculator (an instrument notoriously bad for calculating large values), the following two illustrations of the Earth's rotation gives a general idea of how much the Earth has slowed, and the adjustments needed to be made during the course of developing the following table.
 The rate of rotation today. 1000 MPH at equator 1 day =23 hrs, 56 min., 4.09 sec. |
 The rate of rotation billions of years ago. 3000 MPH at equator approx. 8 hour day |
| Years Ago | Epoch | Period/Age | Era | Eon | Major Events | approx. rotation rate \value used) |
|
|---|---|---|---|---|---|---|---|
| Present day | Holocene | Quaternary | Cenozoic | Phanerozoic |
23.93 hours \.0000001 |
||
| 11,430 (The person in the "Man From Earth" movie is about 3,000 years older than this.) | Pleistocene | Evolution of fully modern humans. Extinction of many large mammals. |
23.934 hrs \.0000001 |
||||
| 1.81 million | Pliocene | Tertiary | Neogene | 23.8977 \.0000002 |
|||
| 5.33 million | Miocene | 23.969 hrs/ 341.28 days \.0005 |
|||||
| 23.0 million | Oligocene | Paleogene | 23.866 hrs/ 389.116 days \.0005 |
||||
| 37.2 million | Eocene | Appearance of first "modern" mammals. | 384.465 days 22.8 hrs |
||||
| 55.8 million | Paleocene | 22.6 hrs | |||||
| 65.5 million | Cretaceous | Mesozoic | Dinosaurs reach peak, become extinct. Primitive placental mammals. |
22.47 hrs/ 342.770 days \.0009 |
|||
| 146 million | Jurassic | Marsupial mammals, first birds, first flowering plants. | 22.4532 hrs | ||||
| 200 million | Triassic | First dinosaurs, Egg-laying mammals. | 19.87 hrs | ||||
| 251 million | Permian | Paleozoic | Permian extinction event- 95% of life on Earth becomes extinct. | 18.84 hrs | |||
| 299 million | Carboniferous | Pennsylvanian | Abundant insects, first reptiles, coal forests. | 17.86 hrs | |||
| 318 million | Mississippian | Large primitive trees | 17.48 hrs | ||||
| 359 million | Devonian | First amphibians, clubmosses and horsetails appear, progymnosperms (first seed bearing plants) appear. | 20.2922 hrs \.0000001 |
||||
| 416 million | Silurian | First land plant fossils. | |||||
| 443 million | Ordovician | Invertebrates dominant. | |||||
| 488 million | Cambrian | Major diversification of life in the Cambrian explosion. | |||||
| 542 million | Ediacaran | Neoproterozoic | Proterozoic | First multi-celled animals. | |||
| 600 million | Cryogenian | Possible snowball Earth period. | |||||
| 850 million | Tonian | ||||||
| 1.0 billion | Stennian | Mesoproterozoic | |||||
| 1.2 billion | Ectasian | ||||||
| 1.4 billion | Calymmian | ||||||
| 1.6 billion | Statherian | Paleoproterozoic | First Complex single-celled life. | ||||
| 1.8 billion | Orosirian | ||||||
| 2.05 billion | Rhyacian | ||||||
| 2.3 billion | Siderian | ||||||
| 2.5 billion | Neoarchean | Archaean | |||||
| 2.8 billion | Mesoarchean | ||||||
| 3.2 billion | Paleoarchean | ||||||
| 3.6 billion | Eoarchean | Simple single-celled life. | 7.333 hrs \.0004 |
||||
| 3.8 billion | Hadean | 20.0791 hrs \.0000001 |
|||||
| 4.1 billion | Oldest known rock. | ||||||
| 4.4 billion | Oldest known mineral. | ||||||
| 4.57 billion | Formation of Earth. | 2.842hours \.0004 |
|||||
Source: Rotation Rate Specific page 2
While the foregoing was no flight of fancy thinking, we must now turn our minds in the direction of being a tad more divergent... If we view differences in electromagnetic radiation as parallel, divergent or some other form of evolution, we could denote them as different species or similar species occurring in a given (atomic-related) environment. In other words, we are animating them and the particles of quantum mechanics. This does not imply sentience or that their is even a brain, but it is like a child bringing paper or other dolls or puppets to life... just as some researchers imaginatively do when working out probable scenarios of a given fossil. It is a qualitative experience we entertain while dreaming... since dreams are not real... our brains give them animation because the guards of the conscious mind are relaxed. Imagination is permitted to run free in dreams (though many do not care to provide details thereof), and may at first seem rather odd if encountered when someone is discussing information related to "serious" topics, but it remains nonetheless useful; whether or not some readers are comfortable with it (at least in a waking state where they have got to keep up their guard of proper appearances in what they write, say, or experimentally construct). It is disturbing for some to "dream out-loud" beyond certain conventions such as "what would you do if you won the lottery?" or some other make-believe musing shared amongst those who think you're nuts anyway... and will let you have your say because they typically lead to some amusement they can join in.
finding parallels, divergences, convergences, etc., amongst different subjects is a important prerequisite in detailing universality or individualized "speciation" of an idea, experiment, application, etc... In other words, for the context at hand, we are looking elsewhere in our analysis of the Peace/War dichotomy in order to identify whether it is a specificity of mind in its current structure, or is the "Peace/War" labeling more on the order of finger-painting that the mind of humans engages in... and many find it of so much interest they give it great importance, like calling an abstraction a work of art? Is the Peace/War dichotomy just another duality amongst others which reflect a two-patterned partitioning that they become obsessively inclined to repeat in the same or some other form?
Date of Origination: Sunday, 22-Jan-2017... 9:52 AM
Date of initial posting: Saturday, 28-Jan-2017... 1:21 PM
Updated posting: Saturday, 31-March-2018... 12:07 PM